This page is an appendix to an article in the magazine Clarinet and Saxophone that describes recent work on articulation from our lab. This page appends sound and video files; it also links to the scientific papers that have more detailed information.
For more background information, see our Introduction to clarinet acoustics or (another non-technical article) The acoustics of woodwind instruments or our multimedia background to waves and sound.
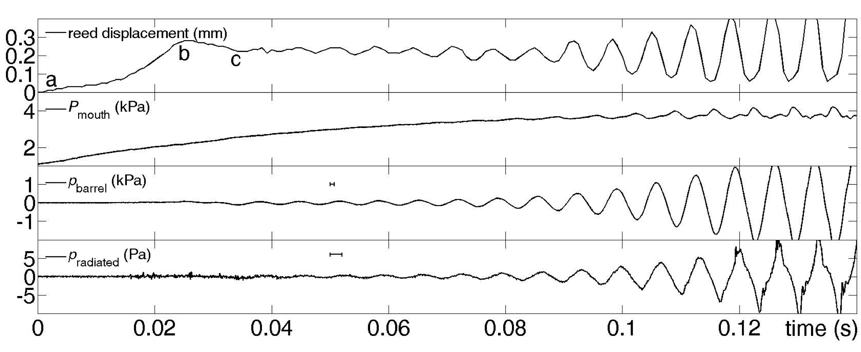
We begin with a measurement from Inwood et al. (2016).
Here, tonguing by players was studied using a mouthpiece on which a pressure transducer was mounted, to measure the blowing pressure. Microphones in the barrel and near the bell measured sound. An endoscope mounted next to the mouthpiece allowed high speed video of the player's tongue and also (when the tongue was not in the way) the motion of the reed.
In this example, over about the first 20 ms, the reed is pulled away from the mouthpiece (a-b), beyond its equilibrium position, by the saliva on the tongue. After release at (b), the reed relaxes to its equilibrium position (c), losing the potential and kinetic energy imparted by the tongue. Meanwhile, however, a pressure pulse due to the changing aperture between reed and mouthpiece has travelled down the bore and back. Successive interactions of the pulse with the reed amplify the signal and build up the standing wave in the bore. The note is D3 (written E3), the lowest on the Bb clarinet.
Severally qualitatively different tongue motions were observed among players; this one is chosen for analysis because the reed was most clearly visible. (The first note in the sound file is the one in the graphs. The horizontal bars in the third and fourth graphs show the distance between microphone and reed divided by the speed of sound. Note too that the tongue below the reed can pull the reed open, because of the saliva on the tongue and time required for air to enter the gap between tongue and reed.)
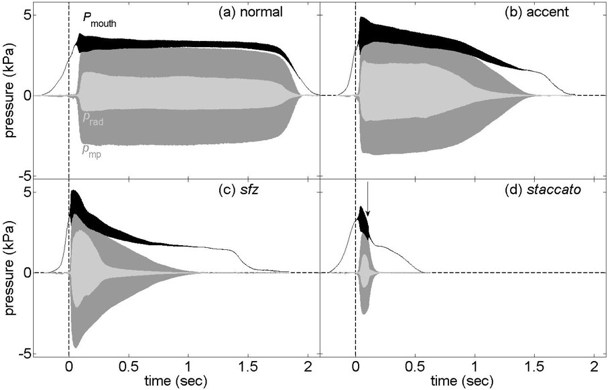
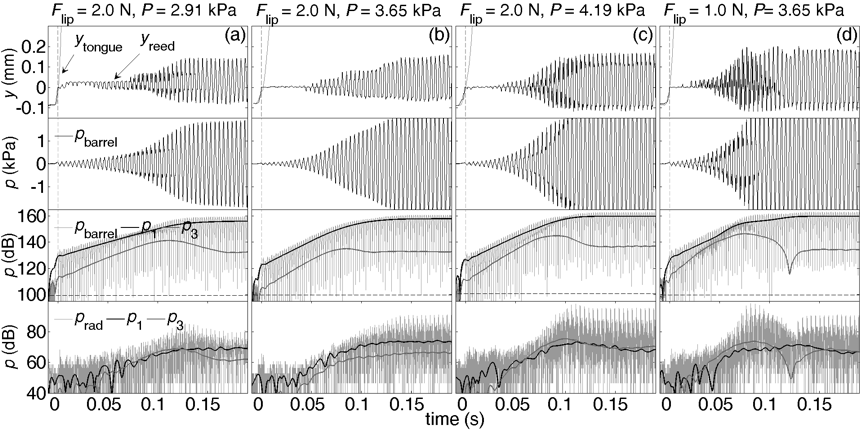
Two figures from another study on human players (Li et al., 2016a) are reproduced here. The pressure measured inside the player's mouth (Pmouth) is plotted in black. (This includes the sound pressure, so the black line is thicker when the note is playing.) The pressure inside the mouthpiece (pmp) is dark grey and the radiated sound near the bell (prad) is light grey. Only Pmouth has a substantial DC component. Below them are two sound files for each graph: one was recorded inside the barrel of the clarinet, near the mouthpiece; Immediately below this is the sound recorded simultaneously near the bell. In the sound file, the barrel recording is the darker timbre, because it is dominated by the fundamental, whereas the high harmonics radiate more efficiently from the bell. (This difference in harmonic content is also visible in the graph in the preceding section, where the shape of the bore signal is more sinusoidal.)
The note is written C5 (sounding Bb4 on this transposing instrument). On these graphs, t = 0 (vertical dashed line) is the instant when the tongue ceased contact with the reed. In the first figure, four different articulations are shown. For the normal articulation, the player played the note given in standard notation with only the expression mark mf (moderately loud). For the accent, '>' was marked over the note. As is usual in music, sforzando was indicated with the letters sfz and staccato with dots above the notes. In the staccato graph, the vertical arrow shows the moment when the tongue touched the reed to stop the note; the tongue was not used to stop the note in any other articulation.
barrel
radiated
normal
accent
barrel
radiated
sforzando
staccato
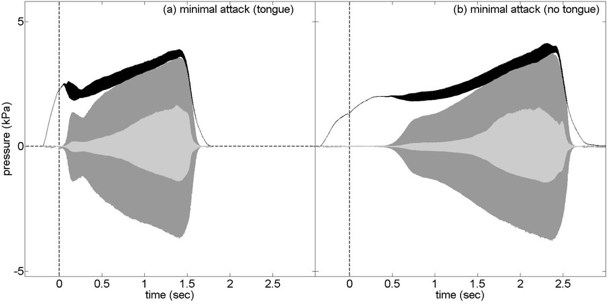
In the following two graphs, the same player was asked to start the same note 'as softly as possible', both using the tongue (left) and without using it.
barrel
radiated
with tongue
without tongue
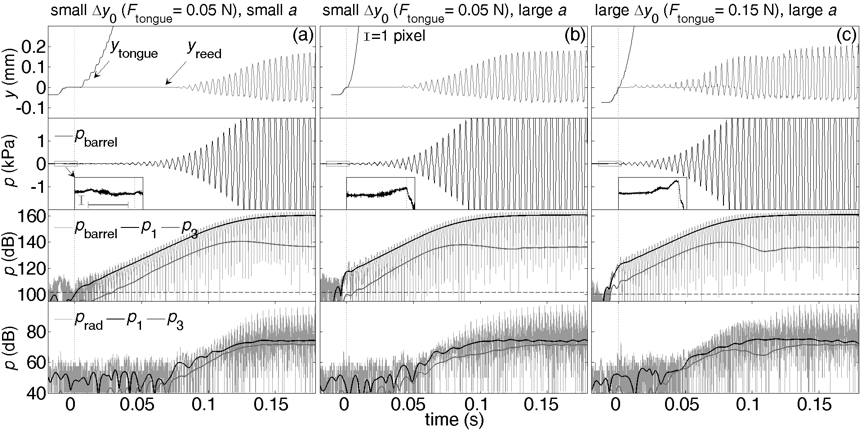
Effect of initial reed displacement and tongue acceleration
Figures from the paper using a clarinet-playing machine are reproduced here. (The clarinet-playing machine itself is on this page.) Below each figure are sound files for each graph: the top row is the sound recorded inside the barrel of the clarinet, and below that is the radiated sound measured near the bell. The barrel recording has a darker timbre, because it is dominated by the fundamental, whereas the high harmonics are radiated more efficiently by the bell, giving a brighter sound.
The top row in the figure plots the displacement y of the reed and the tongue, this time measured from the position of mechanical equilibrium before tongue release, at the tip of the mouthpiece. Two values of initial reed displacement Δy0 and two values of tongue acceleration a and force Ftongue are compared. The second row is a linear plot of the pressure in the barrel of the clarinet (pbarrel). On the bottom two rows, each of the graphs plots three pressure signals on a logarithmic scale. The oscillating signal in pale grey is, again, pbarrel. The black line is the amplitude p1 of the first harmonic of pbarrel. The grey line is the amplitude p3 of the third harmonic. The magnified insets show the change in pbarrel at tongue release; the two scale bars are 20 Pa and 10 ms, respectively.
Below the sound files are links to reduced versions of the videos used to produce the top row of graphs. These have been undersampled (from 8000 to 100 fps) to keep the files small and the videos short. (The short versions will typically run at ¼ speed).
After tongue release, the timing of the growth in reed vibration and sound depend on the initial displacement y0 of the reed and the tongue acceleration a. Here t200pa is defined as the time taken for pbarrel to reach 200 Pa (140 dB with respect to 20 μPa) after tongue release. The t200pa for (a) to (c) above is 77, 41 and 32 ms, indicating that vibration and sound reach higher values sooner following either a larger initial tongue displacement or a larger tongue acceleration. However, this variation does not appear to be caused by the reed having a greater initial potential energy (due to larger y0) or larger kinetic energy at release (larger a), because in each case the reed stops in its equilibrium position after tongue release for at least about 10 ms. Instead, the effect of y0 and a on the onset appear to be due to their transient effects on the air flow into the mouthpiece: in (c), the initially closed reed channel opens in about 7 milliseconds. p1, the amplitude of the first harmonic in pbarrel, shows a very rapid increase above noise levels that coincides with the action of tongue release in (b) and (c) above. This shows that either large tongue acceleration a or large initial displacement y0 of the reed can produce the immediate large change in p1. (See Almeida et al (2017) for explanation.) Similar sudden rises above noise in the p1 in the barrel were also observed when human players performed accents (see above). In both cases, however, it should be noted that, outside the bell, p1 does not rise suddenly above the noise (bottom row of Fig. 2) and no 'click' is heard in the sound files provided for these figures.
As the vibration of the reed grows, it quickly becomes asymmetrical as the reed approaches the lay. For same tongue force Ft and initial displacement y0 of the reed [(a) and (b)], the main difference in reed motion due to the large change in a is that, for large a, the vibration reaches any given value earlier. When there is both a large initial displacement y0 and a large acceleration a (figure c above), there is (for this example) a further difference: the third harmonic rises abruptly above background and remains proportionally stronger during the rise in both reed motion and sound. In the barrel pressure, the third harmonic is below the first by about 14 dB in (a) and (b) and 12 dB in (c). In the quiescent state, however, the third harmonic has the same level (86 dB) in all three cases. (In the radiated sound, the sound level of the third harmonic is closer to that of the first because the bell radiates high frequencies better than low.)
For the three cases shown above, the exponential rate r of increase of the fundamental in pbarrel is 475±20 dB.s−1, indicating that (with constant P and F), the tongue force and acceleration have little effect on r. The amplitude (RMS) of the third harmonic p3 also shows an exponential increase with similar rate. This similarity in r is an important result: while the action of the tongue affects the timing of the rise, it has little direct effect on the transient. Instead, the tongue motion has an important effect on the air flow, because moving the tongue and reed changes the aperture into the mouthpiece (Almeida et al., 20017).
Effect of lip force and blowing pressure
For the cases in the next figure, the tongue force and tongue acceleration were held constant while the steady force F exerted by the lip and the steady blowing pressure P were varied between experiments. The same variables are plotted as in the figure above. The first three rows show the effect of varying mouth pressure. Comparing the second and fourth rows shows the effect of lip force.
The examples above exhibit initial transients that are initially exponential (and so appear linear on these dB plots). According to the simple model in Li et al (2016b), the rate r of exponential growth in the initial transient is given by
r = (10 log10e)*(Rreed+Rbore)/(RreedRboreC),
where the small signal conductance 1/Rreed = ∂U/∂P is the slope of the quasi-static dependence of the flow U past the reed on the mouth pressure P, C is the compliance in a parallel RboreLC resonance that empirically represents the impedance peak of the bore near which the instrument plays, Rbore represents the losses in the bore.
At high values of P, ∂U/∂P is negative because increasing P tends to close the reed aperture. When Rreed is negative and –Rreed < Rloss, r is positive and the initial transient has an exponential rise which continues until the small signal approximation fails and the defnition of Rreed no longer represents the U(P) curve. For any given lip force F, there is a finite range of the
the U(P) curve that satisfies
–Rreed < Rloss, and this determines the playing régime.
Negative rates of exponential growth
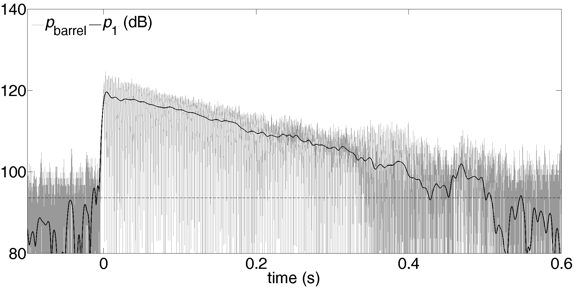
At values of P below the playing range mentioned above, the slope of the U(P) curve is insufficiently negative, so that –Rreed > Rloss, which gives a negative r, so exponential decay is predicted. An example is given below: P is 2.34 kPa and F is 2.5 N. An abrupt increase in pressure is due to the disruption of air flow when the tongue releases the reed. This initiates a standing wave in the bore, whose amplitude decays exponentially, with a rate that may be slower than that produced when the reed is immobilised by the tongue (as at the end of a staccato note).
The sound level is low, especially the radiated sound.
As before, the grey line is the total pressure, the black line the amplitude of the first harmonic.
pbarrel
pradiated
Ending a note
The normal way of ending a note is lowering the blowing pressure to reduce the reed gain so that (as explained above) –Rreed > Rloss, which gives a negative r and a decreasing amplitude. The decay is not quite exponential, because the gain is usually falling slightly during the transient. Changing the bite can also lower the reed gain.
Both methods should give relatively slow decays because the decay starts with the reed at the critical point for a sustained note, and it takes time to vary pressure or bite. Slow pressure variation gives longer decay.
A faster decay is produced by stopping the reed with the tongue, which is what most of our participants did when playing fast staccato notes. In this case, the reed gain falls quickly to near zero. In these cases, the decay rate is determined solely by the losses in the bore. We can determine these losses independently when we measure the acoustic impedance spectrum of the clarinet for a given fingering (without a player) and it's satisfying that the two sets of measurements are consistent.
A note can also be ended by changing Rloss. For instance, relaxing the grip so as to allow leaks under a pad increases the loss. A deliberate change in fingering to one with higher losses can also end the note. These last methods are probably performed more commonly as mistakes than as deliberate strategies.
The tongue need not touch the reed
It's
worth observing that the tongue need not touch and release the reed
to tongue a note. In double tonguing, for example, the 'k' part of the 't-k' alternation uses the tongue to seal the air pathway when it touches the hard palate. It's also possible to tongue using a 'd' action: the tongue seals the air pathway by touching the palate near the root of the front upper teeth, without touching the reed. In these cases, it is the sudden opening of the air pathway that allows a pulse of air with positive pressure to travel
into the instrument and start the 'water hammer' effect, which in turn is amplified by the reed.
The NICTA-UNSW clarinet playing robot. This page also includes the results of a study into how the bite force and mouth pressure influence pitch, loudness and timbre.